
TEMA 5: FISIOLOGÍA DEL ERITROCITO. GRUPOS SANGUÍNEOS
ERITROCITARIOS.
Elementos formes
de la sangre
Existen tres
categorías de elementos formes:
- Eritrocitos, hematíes, células o corpúsculos rojos de la sangre, glóbulos rojos.
- Leucocitos, glóbulos blancos o células blancas de la sangre.
- Plaquetas o trombocitos.
Los eritrocitos; son discos bicóncavos, no
nucleados, con un diámetro medio de 8.5 micrómetros, un espesor en los bordes
de 2 micrómetros y un espesor en el centro de un micrómetro. Esta forma de disco
bicóncavo representa la superficie máxima de difusión para los gases en función
de su tamaño y además esta forma lo hace muy flexible.
Carecen de núcleo,
sin embargo consumen glucosa, oxígeno y ATP y liberan CO2. Esto
quiere decir que realizan procesos metabólicos que emplea para alimentar los
sistemas de transporte activo que mantiene la homeostasia iónica entre la
célula y el medio, en este caso el plasma.
El número varía
entre los 4,5 a 6 millones / mm3 en el varón a los 4 o 5,5 millones
/ mm3 en la mujer. El número también varía con la edad.
HEMATOPOYESIS
Este término se utiliza para designar la formación
de los elementos formes de la sangre, pero su mecanismo de formación no es
homogéneo y hay que distinguir los elementos del tejido linfoide, de los
elementos del tejido mieloide, pro sin embargo, los elementos de estos dos
tejidos tienen precursores embriológicos comunes y posiblemente en el adulto
siguen existiendo células germinales comunes a ambas líneas. Los dos tejidos
difieren totalmente, así, el tejido linfoide dará origen a un tipo de glóbulo
blanco que son los linfocitos y por el contrario desde el tejido mieloide se
van a originar el resto de elementos formes de la sangre.
Los distintos
elementos de este tejido mieloide tienen numerosos puntos en común, ya que se
producen a partir de precursores que es la unidad formadora de colonias del
bazo, y de hecho en el adulto, el lugar exclusivo de origen de médula
hematopoyética es la médula ósea, en donde todos los elementos formes de la
sangre sufren dos tipos de modificaciones; se multiplican y se diferencian.
El lugar de
formación de las células mieloides es diferente en el embrión, en el feto y
tras el nacimiento. Así, durante la vida intrauterina el lugar de formación de
estas células es el saco vitelino, el bazo, el hígado, los nódulos linfáticos y
la médula ósea roja. En el momento del nacimiento, e incluso un poco antes, de
todas las zonas, excepto la médula ósea roja se vuelven improductivas para
células mieloides. Esta médula ósea roja se halla en el esternón, en las
costillas, en las vértebras, en los huesos del cráneo y en los extremos de los
huesos largos, por lo tanto vamos a hablar de mielopoyesis, que es el proceso de formación de elementos formes de
la sangre a partir de células mieloides.
En la médula ósea
roja hay unas células madre hematopoyéticas pluripotenciales, de las cuales
derivan todas las células de la sangre circular, una porción de estas células
permanece igual durante toda nuestra vida, y otra porción mayor se diferencia.
La primera
descendencia no podemos diferenciarla morfológicamente desde ningún punto de
vista de la anterior, sin embargo si la hacemos crecer, veremos que ya está
comprometida con una línea que es la línea mieloide y la línea linfoide. Por lo
tanto, a esta primera descendencia se le llama células madres comprometidas o
prediferenciadas y son dos:
- La unidad formadora de colonias del bazo.
- La célula madre linfoide.
De la unidad
formadora de colonias del bazo se originan tres tipos de células:
Ø La unidad formadora de colonias blásticas que luego se vuelve a
diferenciar en la unidad formadora de colonias de eritrocitos y que dará lugar
a los eritrocitos.
Ø La unidad formadora de colonias de granulocitos y monolitos, que dará
lugar a todos los glóbulos blancos excepto los linfocitos.
Ø La unidad formadora de megacariocitos, cuyo megacariocito al romperse
da lugar a las plaquetas.
Unidad formadora de Células del Bazo:
- Unidad formadora de células blásticas Þ unidad formadora de células de eritrocitos.
- Unidad formadora de granulocitos y monolitos Þ Glóbulos blancos.
- Unidad formadora de megacariocitos Þ Plaquetas.
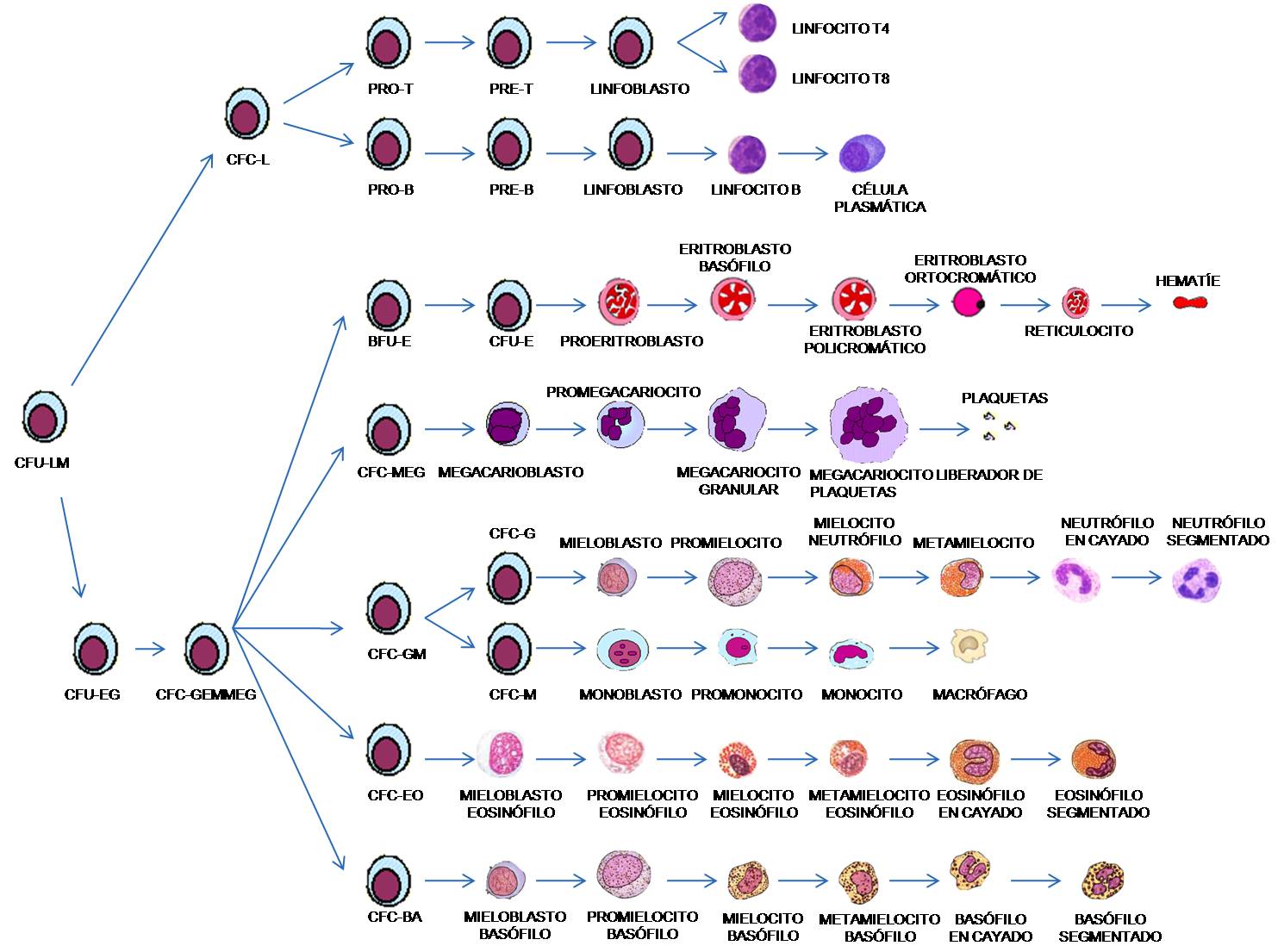
ERITROPOYESIS
Es el proceso de
formación de eritrocitos. Los glóbulos rojos se forman en varias zonas de
nuestro cuerpo, dependiendo si estamos en fase letal o después del nacimiento;
- En el primer trimestre de la gestación los glóbulos rojos se originan en el saco vitelino, esta etapa se llama mesoblástica y el glóbulo rojo que se produce es de gran tamaño y se llama megalocito.
- En el segundo trimestre los glóbulos rojos se producen en el hígado, esta etapa se llama hepática y el glóbulo rojo que se produce ya no posee núcleo y se le llama normocito.
- En el tercer trimestre se produce ya en la médula ósea roja, por lo que esta etapa se llama mieloide y el glóbulo rojo ya es el eritrocito. Esta etapa se prolonga hasta los 20 años de edad pero a partir de esta edad la médula ósea de huesos largos se vuelve improductiva, y por lo tanto los glóbulos rojos se producen en la médula ósea roja de los huesos membranosos.
- A medida que avanzamos en edad, la médula ósea roja se vuelve menos productiva, de ahí que muchas personas mayores puedan presentar anemias. El organismo, en estos casos estimula de nuevo al hígado y al bazo para que vuelvan a producir glóbulos rojos.
En la médula ósea
roja el productor de eritrocitos es la unidad de formación de colonias de
eritrocitos, este es un precursor, a partir del cual, mediante división y
diferenciación se producen proeritroblastos, que son la primera célula que
podemos reconocer de esta línea, el proeritroblasto se divide y se diferencia y
da lugar al eritroblasto I o eritroblasto basófilo en donde comienza la
síntesis de hemoglobina. El eritroblasto I se divide y se transforma en
eritroblasto II que también es basófilo (es decir, posee núcleo). El eritroblasto
II se vuelve a dividir y diferenciar y se convierte en policromatófilo, a
partir de aquí el eritroblasto policromatófilo sufre un proceso de
diferenciación y se convierte en el eritroblasto reidófilo, también llamado
normoblasto.
Este normoblasto se
vuelve a diferenciar y se transforma en reticulocito, que no tiene núcleo, sólo
filamentos de cromatina, y tiene gran cantidad de hemoglobina. Hasta este
momento todo se ha producido en la médula ósea, luego los reticulocitos salen a
la sangre, pierde totalmente lo que les queda de núcleo y se transforman en
eritrocitos, y de hecho podemos encontrar en nuestra sangre una pequeña
cantidad ( entre el 0,5 – 1%) de reticulocitos circulando en sangre.

Las sustancias
necesarias para que se produzca la eritropoyesis son;
- Lípidos, fundamentalmente fosfolípidos y colesterol, utilizados para incorporarse a la membrana y al estroma.
- Proteínas utilizadas fundamentalmente para las membranas.
- Hierro, se necesita para la síntesis de hemoglobina.
- Aminoácidos, se necesitan para la síntesis de hemoglobina.
- Eritropoyetina, es un enzima glucoproteína que es liberada por el riñón cuando es necesaria una eritropoyesis, necesario para estimular la producción de eritrocitos.
- Vitamina B12, se usa en la formación de DNA en la maduración nuclear.
- Factor intrínseco, es un mucopolisacárido segregado por el estómago que tiene la misión de combinarse con la vitamina B12 y asegurar la absorción de esta vitamina a nivel intestinal.
- Piridoxina, es una sustancia que aumenta la velocidad de división celular.
- Cobre, cataliza la formación de hemoglobina.
- Cobalto, cataliza la formación de hemoglobina.
- Ácido fólico, promueve la síntesis de DNA en la división celular.
Cuando las
eritropoyesis son normales y eficaces, la médula ósea roja produce
aproximadamente 3.5 millones de células maduras por Kg. de peso corporal y día.
Estas células circulan entre 90 y 120 días realizando las funciones de
transporte de O2 y CO2. Al final de estos 120 días los
eritrocitos viejos son fagocitados por células del sistema retículo endotelial
localizados fundamentalmente en hígado, bazo y médula ósea, pero estas células
fagocitan también a todo glóbulo rojo anormal. Reconocen las células viejas y
anormales por cambios químicos que se producen en las membranas de estas
células. Parte de los materiales fagocitados van a ser reutilizados para una
nueva síntesis de hemoglobina y eritrocitos
o de de otras células. Uno de los componentes esenciales del glóbulo
rojo es la hemoglobina.
Metabolismo de la Hemoglobina
La cantidad de
hemoglobina es de 34g /100 ml de glóbulos rojos, lo que quiere decir que en el
hombre existen 16g de hemoglobina/100 ml de sangre y en la mujer 14 g de
hemoglobina/100 ml de sangre. La hemoglobina es el pigmento respiratorio de
vertebrados. Está formada por 4 núcleos, cada uno de los cuales se llaman
grupos Hemo o grupos Hem y poseen 4 anillos y un átomo de hierro. Además la
hemoglobina posee unas cadenas polipeptídicas que en su conjunto denominamos
globina.
La hemoglobina
posee hierro, el cual está unido a los cuatro nitrógenos de los anillos y que
además se une a la globina. Cuando la hemoglobina transporta oxígeno, el hierro
se une a éste, formándose en este caso un compuesto llamado oxihemoglobina o
hemoglobina oxigenada.
La hemoglobina
cuando transporta el oxígeno no se oxida, sólo se oxigena, ya que el hierro
permanece ene estado ferroso, por esto, la envían del oxígeno con el hierro es
una unión muy fácil de romper y reversible. La estructura macroscópica de la
hemoglobina es;
Síntesis de Hemoglobina
Empieza a sintetizarse
en la fase de eritroblasto, pero la mayoría de sus componentes tienen origen
hepático. La síntesis de hemoglobina proviene del ácido acético y de la
glicina. Lo que ocurre en su síntesis es;
- El ácido acético en el Ciclo de Krebs se convierte en Succinil – CoA.
- Dos Succinil – CoA se unen con dos glicinas y dan un anillo pirrólico, un pirrol.
- Cuatro pirroles se unen y forman la protoporfirina que en el hombre es la protoporfirina IX.
- Esta protorfirina IX se une con el Fe y forma el grupo hemo.
- Un grupo hemo se une con una cadena polipeptídica que puede ser α o β y nos da la cadena de hemoglobina α o la cadena de hemoglobina β.
- dos cadenas de hemoglobina α se unen con dos cadenas de hemoglobina β y tenemos la hemoglobina A.
Por lo tanto, la
globina está formada por cuatro cadenas polipeptídicas, la más común es la que
está formada por dos cadenas α y dos cadenas β; las α poseen cada una 141
aminoácidos y las β 146 aminoácidos. La alteración simple de la secuencia de
aminoácidos en alguna de estas cadenas determina que esa hemoglobina no sea la
hemoglobina normal. En la etapa fetal se produce otro tipo de hemoglobina que
es la hemoglobina fetal, que no tiene cadenas β, sino que tiene unas cadenas
que denominamos cadenas γ. Esta hemoglobina es muy ventajosa para la especie
humana porque es esta hemoglobina la que tiene una mayor capacidad de captación
del oxígeno, por lo que capta el oxígeno a presiones de oxígeno en sangre más
bajas. La presión parcial del oxígeno en sangre fetal es más baja que la
presión parcial de oxígeno en sangre arterial de un adulto.
Metabolismo del Hierro
El hierro es
esencial para el transporte de oxígeno, se absorbe en la primera parte del
intestino delgado (duodeno), esta absorción es por transporte activo. Su
absorción es más rápida en estado ferroso que férrico y también es más fácil en
forma de hierro hemínico que en no hemínico, por lo que se absorbe mejor el
hierro de carnes rojas que el hierro de vegetales.
Esta absorción se
debe a que el hígado segrega en la bilis cantidades pequeñas de una proteína
transportadora que es la apotransferrina. Esta proteína a través de la bilis va
al duodeno donde se une a los receptores de membrana de las células epiteliales
de la pared intestinal y así transporta el hiero pasando esta transferían al
torrente sanguíneo.
El hierro absorbido
o el hierro que existe en la sangre producto de la destrucción de los
eritrocitos va a estar combinado con la apotransferrina en forma de
transferían, y así va a circular por la sangre para ser llevada a los lugares
de utilización o de depósito, de hecho, el hierro en exceso presente en la
sangre se va a depositar en todas las células del organismo, pero especialmente
en las células hepáticas, donde se va almacenar el 60% de hierro en exceso.
En el hígado el
hierro se libera de la apotransferrina y se combina con otra proteína que es la
apoferritina formando un compuesto que es la ferritina o hierro en depósito.
Esta ferritina va liberar hierro cuando es necesario.
El hierro puede
almacenarse también de forma más insoluble en un compuesto llamado hemosiderina
que forma grandes acumulos en las células, al ser más insoluble su
almacenamiento sólo ocurre cuando la cantidad de hierro en el organismo es
mayor que el que puede contener la apoferritina.
El hierro también
va a ser utilizado para la síntesis de hemoglobina. Este hierro es llevado a la
médula ósea en forma de transferían y esta transferían se fija en los
receptores de membrana de los
eritroblastos que le van a ingerir por endocitosis y una vez en el interior la
transferían descarga el hierro a nivel mitocondrial en donde se va a producir
la síntesis de los grupos hemo y luego vuelve a salir en forma de
apotransferrina al plasma.
La pérdida de
hierro en nuestro organismo es de dos tipos:
- A través de la orina (se pierde 0,6 mg/día) y a través de las heces.
- Pérdida de hierro por pérdida sanguínea, lo normal es que el varón no tenga esta pérdida de hierro, pero en la mujer representa una pérdida de 0,7 mg/día de hierro durante los días de la menstruación.
Proceso de destrucción de los glóbulos rojos
Al romperse un glóbulo
rojo, la hemoglobina pasa al torrente circulatorio y va a sufrir una serie de
transformaciones;
- Toda hemoglobina va a ser fagocitada por macrófagos en distintas partes de nuestro cuerpo, pero fundamentalmente por los macrófagos del hígado (que se llaman células de Kopffer) por macrófagos del bazo y por macrófagos de la médula ósea.
- Allí en los macrófagos a las horas o días siguientes se produce la liberación de globina que va a ser reutilizada y la liberación de hierro, que también va a ser reutilizado.
- Lo que queda es la porción porfirínica y esta porción en el interior del macrófago sufre varias transformaciones, convirtiéndose en un pigmento, que luego va a ser un pigmento biliar llamado bilirrubina.
- Esta bilirrubina va pasar a la sangre y va a ser transportada al hígado donde va a sufrir una serie de transformaciones.
Formación de la Bilirrubina
A nivel de los
macrófagos la hemoglobina se desdobla debido a un enzima hemo – α –
metanooxigenasa en globina y grupos hemo, el grupo hemo se abre y pierde el
hierro, entonces nos quedan unas cadenas rectas de grupos pirrólicos que es la
porción porfirínica. Esta porción porfirínica se transforma en un compuesto de
color verde que es la biliverdina, que debido a un enzima que es la biliverdina
reductasa se transforma en bilirrubina que va a ser liberada del macrófago de
forma gradual hacia la sangre.
Esta bilirrubina
libre en el plasma se combina con mucha rapidez con una proteína transportadora
que es la albúmina y de esta forma va a circular por el plasma y el líquido
intersticial, pero en el plasma puede existir también hemoglobina libre
producto de las hemólisis intravascular de los hematíes.
Esta hemoglobina
libre tiene que ser captada por unas proteínas que tienen como misión principal
formar complejos con la hemoglobina y transportarla hacia las células del
organismo, fundamentalmente las hepáticas y allí se transformará también en
bilirrubina.
Estas proteínas
transportadoras son las haptoglobinas que fijan dímeros de hemoglobina, la
hemopexina, que fija los grupos hemo y la albúmina que fija también hemos
oxidados.
En plazo de horas,
la bilirrubina formada por un camino u otro y transportada por la albúmina va a
ser absorbida por las membranas de las células hepáticas y allí se va a liberar
de la albúmina y se va a unir a dos tipos de proteínas dentro de las células
hepáticas (hepatocitos). A continuación y a este nivel la bilirrubina se
conjuga con ácido glucurónico o con sulfato, y una vez que se ha formado la
bilirrubina conjugada y de una manera lenta pasa una pequeña cantidad al plasma
de nuevo, y la mayor parte pasa al intestino a través del jugo biliar o bilis.
Una vez en el intestino esta bilirrubina conjugada, por acción bacteriana se
convierte en urobilinógeno y este urobilinógeno sigue dos caminos, una parte va
a ser absorbida por la pared intestinal y pasa al plasma y desde allí, en el
proceso de filtración a nivel glomerular va a pasar a la orina de donde va a
ser eliminado, pero este urobilinógeno cuando se pone en contacto con el aire
se oxida y se transforma en urobilina que es la forma en que encontramos el
urobilinógeno en orina.
La otra parte que
permanecía en el intestino va a ser eliminada a través de las heces, pero el
contacto con la sangre se transforma en urobilina, que en este caso reciben los
siguientes nombres:
Uribilinógeno = Estecobilinógeno (¿mirar este nombre?)
Urobilina = Estecobilina
Una pequeña
cantidad de la bilirrubina conjugada que había pasado al plasma va a ser
eliminada también a través del riñón en forma de bilirrubina conjugada.
Funciones de los Glóbulos Rojos
- Transporte de oxígeno y dióxido de carbono, de hecho, un gramo de hemoglobina es capaz de transportar 1,33 cm3 de oxígeno.
- Pueden actuar como elementos o sistemas tampón, es decir, amortiguadores, porque poseen un enzima que es la anhidrasa carbónica que cataliza la siguiente reacción:
Y por lo tanto en la sangre puede actuar con una gran cantidad de CO2.
Alteraciones en el número de glóbulos rojos
La disminución en
el número de glóbulos rojos se conoce con el nombre de anemia, que puede ser
producida por varias causas.
La primera
consecuencia de anemia es un déficit en el transporte de oxígeno, pero también
produce una disminución de la viscosidad, por lo tanto la sangre se vuelve más
fluida, por lo que circulará con mayor rapidez y el corazón tendrá que trabajar
más. En consecuencia, si un estado anémico persiste se puede presentar
insuficiencia cardiaca.
El aumento del
número de glóbulos rojos se llama policitemia, por lo que la viscosidad está
aumentada, el fluido sanguíneo es más lento, pero el corazón necesita trabajar
más para mover algo más viscoso, por lo tanto, si una policitemia persiste se
presentará también insuficiencia cardiaca.
Grupos Sanguíneos Eritrocitarios
Vamos a ver
solamente los dos sistemas eritrocitarios más estudiados, que son los sistemas
A, B, 0 y el sistema Rh.
SISTEMA A, B, 0
Bordet puso en 1895
de manifiesto que al mezclar sangre de dos especies diferentes se produciría un
fenómeno de aglutinación que se llamó heteroaglutinación.
Un poco más tarde,
en 1900, Landstener puso de manifiesto que este fenómeno de aglutinación
ocurriría muchas veces al mezclar sangre de la misma especie y llamó a este
nuevo fenómeno isohemoaglutinación, por lo que se llegó a la conclusión que en
los glóbulos rojos existían unos mucopolisacáridos, los aglutinógenos que
actúan como antígenos y que en el suero existían otros mucopolisacáridos, las
aglutininas, que actuaban como anticuerpos y por lo tanto al mezclar los
complementarios era cuando se producía el fenómeno de aglutinación, que no era
ni más ni menos que una reacción inmunitaria.
Se dijo que
existían 4 tipos sanguíneos para este sistema;
TIPOS
|
Glóbulos Rojos
(Aglutinógenos)
|
Suero
(Aglutininas)
|
A
|
A
|
Anti – B
|
B
|
B
|
Anti – A
|
AB
|
A y B
|
|
0
|
Anti – A y Anti - B
|
Según esto, al cero
se le llamó dador universal, porque no tenía aglutinógenos, pero no es
totalmente cierto porque tiene aglutininas y va a producir aglutinaciones con otros
tipos, pero estas aglutinaciones son muy pequeñas y no tienen importancia
médica, ya que el volumen introducido es una transfusión va a ser siempre muy
pequeño comparado al volumen circular.
Al AB se le llamó
receptor universal porque no tenía aglutininas, pero pasa lo mismo que en el
caso anterior.
La herencia de
este sistema
Existe la teoría de
Bernstein que dice que existen tres genes (A, B y 0), pero que se comportan
como alelos múltiples de un mismo locus del par de cromosomas homólogos, de tal
modo que A y B son dominantes sobre 0 y codominantes entre sí, según esto
existen 4 fenotipos y 6 genotipos:
Fenotipos
|
Genotipos
|
A
|
AA
A0
|
B
|
BB
B0
|
AB
|
AB
|
0
|
00
|
Al estudiar más
profundamente este sistema se vio que el aglutinógeno A era inestable y que estaba
formado por dos tipos que eran A1 y A2, de tal modo que A1,
A2 y B son dominantes sobre 0, A1 dominante sobre A2,
A1 codominante con B y A2 también codominante con B.
Según esto hay 6 fenotipos y 10 genotipos.
Fenotipos
|
Genotipos
|
A1
|
A1 A2
A1 A1
A1 O
|
A2
|
A2 A2
A2 0
|
B
|
BB
B0
|
A1B
|
A1B
|
A2B
|
A2B
|
0
|
00
|
SISTEMA Rh
Lansteiner
conjuntamente con Wiener pusieron de manifiesto que al mezclar sangre de Macacus
rhesus y conejo, el conejo producía unas aglutininas capaces de aglutinar la
sangre del macaco y frecuentemente del hombre; luego se hicieron estudios y se
vio que en el 85% de los hombres blancos se producía aglutinación y en el 15%
restante no lo había. Entonces pensaron que existían dos tipos paras el factor
rhesus, por un lado los Rh+ y por otro los Rh-, que serían los que no
presentaban aglutinación. Además se comprobó que este sistema era responsable
de la sensibilización que presentaban los pacientes que habían tenido
transfusiones repetidas y en los cuales se había tenido en cuenta el sistema
AB0, y también se comprobó que este sistema era responsable de la
sensibilización madre-hijo.
Se vio que este
sistema se hereda independientemente del sistema AB0, y de otros sistemas
sexológicos y que aparentemente se comportaba como un factor genético mendeliano
sencillo, que estaría regulado por un par genético, de tal modo que el Rh+
sería dominante y el Rh- sería recesivo, pero poco después se puso en evidencia
que el sistema Rh era el complejo de varios factores y se aislaron numerosos
aglutinógenos y la cosa se complicaba todavía más ya que los aglutinógenos
podían estar todos juntos, aislados o faltar algunos.
Es entonces cuando
se formulan varias teorías genéticas sobre él. Dos de las cuales son
consideradas válidas. La más aceptada es la de dos investigadores Fischer y
Race, que admite la existencia de tres pares genéticos (Cc, Dd, Ee), que están
situados en tres locus distintos de un mismo cromosoma, pero muy próximos entre
sí, debido a lo cual la posibilidad de entrecruzamiento es mínima.
“Existen 3 genes (C, D, E), estrechamente ligados, cada
uno de ellos tiene dos o más formas alélicas, por lo tanto al menos tendremos 6
alelos”.
Si tenemos 6
alelos, a nivel de un cromosoma podemos obtener 8 combinaciones posibles de
estos alelos. A estas combinaciones para un carácter a nivel de un cromosoma se
les llama haplotipos, las 8 combinaciones son;
CDe
cDe
cDE
CDE
Cde
cde
cdE
CdE
Si tenemos 8
haplotipos tenemos 64 genotipos, pero de estos 64, sólo se pueden fenotipar 18,
porque fenotípicamente son iguales y porque somos incapaces de distinguir Dd de
DD, ya que d es ausencia y D es dominio sobre d.
La otra teoría es
la de Wiener, quien dice que sólo existe un locus y con una serie muy numerosa
de alelos y que estos alelos son combinaciones de los alelos que presentaba
Fischer, por lo tanto, un solo locus con una serie muy numerosa de alelos, por
lo tanto se hizo una correspondencia entre los haplotipos de Fischer y de
Wiener, con R para todo lo que lleve D y r para todo haplotipo que lleve d.
F – R
|
W
|
Cde
|
R1
|
cDe
|
R0
|
cDE
|
R2
|
CDE
|
R z
|
Cde
|
r’
|
Cde
|
r
|
cdE
|
r’’
|
CdE
|
r y
|
El carácter + o –
que es aquello que se analiza viene sólo determinado por el par de alelos Dd,
de tal modo que serán + los individuos DD y Dd y serán – los individuos dd.
En el laboratorio
se analiza normalmente utilizando sólo el suero correspondiente al aglutinógeno
que queremos probar. Como el aglutinógeno D es el que caracteriza el carácter +
o -, generalmente sólo se utiliza anti-D; pero si queremos analizar realmente
el sistema sanguíneo Rh tenemos que utilizar los 5 antisueros correspondientes:
Anti – D
|
Anti – Rho
|
Anti – C
|
Anti – rh’
|
Anti – E
|
Anti – rh’’
|
Anti – c
|
Anti – hr’
|
Anti – e
|
Anti – hr’’
|
El sistema Rh es el
responsable de la sensibilización madre – hijo y se presenta cuando la madre es
Rh- y el hijo es Rh+, ocurre porque aglutinógenos del hijo pasan a la madre,
entonces ésta se sensibiliza, formando las aglutinas correspondientes y por lo
tanto, cuando esto ocurre se presenta la reacción inmunitaria antígeno –
anticuerpo, y como la reacción inmunitaria es en 2 tiempos, de ahí que el
primer embarazo sea menos arriesgado que los siguientes. Sólo cuando la madre
ya está sensibilizada se produce la reacción que recibe el nombre de
eritroblastosis fetal o enfermedad hemolítica del recién nacido.
Esta
incompatibilidad puede presentarse también para los factores C y E y para el
sistema AB0 aunque es muy raro y poco grave, también puede presentarse por
incompatibilidad para otros sistemas sanguíneos como son el sistema Kell, el
Duffy, el Lutheran y el Kidd.
Importancia de los grupos sanguíneos en las transfusiones
Tienen gran
importancia, ya que si no se transfunde la sangre del mismo grupo al del
receptor se produce la aglutinación de los glóbulos rojos del receptor.
Antes de toda transfusión
se tiene que determinar el grupo de sangre del donante y del receptor, y además
realizar la prueba cruzada. Esta prueba nos va a decir si las dos sangres son
compatibles, y no sólo para los sistemas AB0 y Rh, sino que para todos los
sistemas sanguíneos. En esta prueba se mezclan eritrocitos del donador con
plasma del receptor, y plasma del donador con eritrocitos del receptor y
cualquier mínima evidencia de una reacción antígeno-anticuerpo debe de
llevarnos a rechazar la sangre del donador.
Los líquidos que
podemos transfundir son;
- Sangre total.
- Plasma; esta transfusión se utiliza para mantener el volumen sanguíneo.
- Soluciones coloides como Dextran y albúmina. Cuando transfundimos este tipo de transfusiones hay que controlar el hematocrito.
- Soluciones de cristaloides, éstas se utilizan para aumentar temporalmente el volumen sanguíneo. Sólo temporalmente porque los cristaloides por ser muy pequeños se filtran rápidamente hacia los tejidos.
No hay comentarios:
Publicar un comentario