
Holaaaaaaaaa!!!
Seguimos subiendo temas de "bioquimica". Hoy le toca el turno a la gluconeogenesis . Esa ruta o proceso metabólico que nos permite obtener glucosa a partir de ciertos precursores y por supuesto utilizando la forma de energía denominada ATP ( adenosín trifosfato).
GLUCONEOGÉNESIS
A partir de ellos:
- Piruvato
- Lactato (músculo esquelético)
- Glicerol (tejido adiposo)
- Glutamato (aminoácido) Se puede obtener GLUCOSA en el hígado.
- Aspartato (aminoácido)
- Alanina (aminoácido)
En el tejido adiposo se produce
la movilización de las grasas por una señal nerviosa, y los ácidos grasos serán
llevados a las células hepáticas. La cantidad de lactato generada no es
significativa no puede cubrir las necesidades totales, es necesario el aporte
de piruvato de otras células. Ejemplo: aa que forman parte de proteínas útiles
de otros tejidos. Hay células que pueden ceder mas fácilmente algunas proteínas
como por ejemplo el músculo esquelético, pueden actuar como donador de alanina.
Perdemos peso debido a que perdemos tejido adiposo y masa muscular.
En el hígado a partir de 2 ácidos
pirúvicos se puede obtener glucosa. El 90%
de la gluconeogénesis se lleva a cabo en el hígado, el 10% restante en
la corteza renal. Ningún otro tejido puede sintetizar glucosa. Las células de
tejidos periféricos proporcionan intermediarios y precursores. Los precursores
para la síntesis de glucosa son el piruvato. Las células hepáticas no almacenan
piruvato, este y otros intermediarios tendrán que ser aportados por otras
células.
El ácido Pirúvico que se usa como
sustrato no es el que se había obtenido de la glucolisis. Porque cuando hay
demanda de glucosa es precisamente porque no hay suficiente, se ha degradado
toda con lo que no habrá ácido Pirúvico.
En ayunas los precursores de la
gluconeogénesis son piruvato, lactato (provienen de las células del músculo
esquelético y de los eritrocitos), glicerol (proviene de las grasas formadas
por glicerol + ácidos grasos (estos últimos nunca serán precursores para la
síntesis de glucosa)), aminoácidos (alanina del músculo esquelético. Son
sustratos no glucídicos y por un mecanismo, que no es inverso a la glucolisis,
se produce glucosa.
En este proceso utilizaremos una
ruta anaplerótica en la que se retira Oxalacetato del Ciclo de Krebs para que
sirva de precursor de la glucosa. Mediante la carboxilación de piruvato a
Oxalacetato, pero el Oxalacetato no proviene de la β-oxidación de ácidos
grasos.
En el proceso de formación de la
glucosa muchas ve las reacciones son inversas a las de la degradación. 7 de las
10 etapas son inversas a las de la glucolisis. Las 3 restantes no son inversas,
son las que corresponden a los puntos de control de la glucolisis, son etapas
completamente irreversibles.
No existe senda metabólica alguna
para la formación neta de nueva glucosa a partir de los 2 átomos de C del grupo
acetilo del Acetil-CoA, ni tampoco una mediante la cual los átomos de C de los
ácidos grasos puedan ser utilizados para producir nueva glucosa.
La reacción Piruvato →→→
Fosfoenolpiruvato no es posible ya que es un punto de control, por lo tanto,
irreversible. Sin embargo el piruvato llegará a convertirse en
fosfoenolpiruvato pero mediante una serie de reacciones muy complejas.
El piruvato del medio
citoplasmático tendrá que penetrar en la mitocondria (mediante
transportadores). En la mitocondria va a ser transformado en Oxalacetato.
2
ATP
Piruvato ------------ Oxalacetato. Mediante la Piruvato
Carboxilasa
Acetil-CoA
Piruvato + CO2 + ATP ----------------------- Oxalacetato + ADP + Pi
El propio Acetil-CoA actúa como:
- Modulador negativo de la piruvato deshidrogenasa; Piruvato ----------- X Acetil-CoA
- Modulador positivo de la piruvato carboxilasa; Piruvato ---------------- Oxalacetato
El Oxalacetato una vez que este fuera de la mitocondria por medio de unos intermediarios, se transformará en fosfoenolpiruvato. Como el Oxalacetato no puede salir de la mitocondria, debe transformarse en malato para poder atravesar la doble membrana mitocondrial.
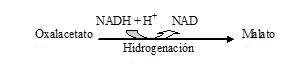
Sale al citoplasma (el malato)
mediante un sistema de lanzadera específico. Una vez allí tiende a experimentar la misma
reacción pero a la inversa regenerándose el Oxalacetato.

De este modo, el poder reductor
cambia de lugar pero no se gasta porque sigue dentro de la célula.
El Oxalacetato también puede
transformarse en Aspartato (aminoácido). El Oxalacetato es un ά cetoácido del
aspartato y captando un grupo amino se transforma en aspartato que puede
atravesar la doble membrana mitocondrial. Una vez fuera, pierde el grupo amino
y se transforma en Oxalacetato.
Hasta aquí el piruvato ha entrado
en la mitocondria, se ha transformado en Oxalacetato y por medio de
intermediarios se ha situado en el citoplasma.
Gasto; 2 ATP debido a las 2
descarboxilaciones.
El Oxalacetato es una molécula
con 4 C y
se puede transformar fácilmente en fosfoenolpiruvato por descarboxilación. La
reacción la cataliza la fosfoenolpiruvato-carboquinasa. El grupo P que hay que
donar para formar el fosfoenolpiruvato es donado por el GTP (GTP → GDP). Se
gastarán 2 GTP porque hay que fosforilar 2 oxalacetatos.
La reacción es;
Oxalacetato (4C ) + GTP -------------------- Fosfoenolpiruvato(3C ) + GDP + CO2
A partir de aquí habrá toda una
serie de intermediarios coincidentes con los de la glucosa, variando los
enzimas.

* 1º punto de control.
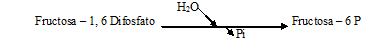
Durante la síntesis de glucosa la reacción;
fructosa -1, 6 DiP + ADP → fructosa – 6P + ATP, se evita mediante una
desviación que se produce gracias al enzima Fructosa-difosfatasa que cataliza
la hidrólisis, esencialmente irreversible, del grupo 1P.
* 2º punto de control.
La producción de glucosa libre es
inducida por la Glucosa – 6 fosfatasa que cataliza la siguiente reacción;

Los dos enzimas se ven
estimulados por la alta concentración de ATP y se ven inhibidos por la baja
concentración de ATP.
Como resultado de la
Gluconeogénesis de obtendrá:
- Glucosa
- 2 NAD+ Þ se habrán gastado; 2 NADH + H+
- 2 GDP Þ se habrán gastado; 2 GTP
- 4 ADP Þ se habrán gastado; 4 ATP
- 6 Pi
En la mayoría de las células la
glucosa – 6P formada durante la gluconeogénesis se emplea como precursor en la
producción de polímeros de reserva (almacenaje) de otros monosacáridos distintos
de la glucosa, de disacáridos y de polímeros estructurales. Sin embargo, en
células como las del hígado, riñón y epitelio intestinal si se puede
transformar la glucosa – 6P en glucosa.
Lo que ocurre es que en las demás
células (músculo, cerebro), no se encuentra la glucosa – 6 fosfatasa por lo que
estos órganos no poseen la capacidad de liberar glucosa libre. Por ejemplo; el
músculo esquelético usa la glucosa – 6P para la vía glucolítica, para consumo
propio, por lo que no requiere glucosa libre.
En
el hígado, ya que existe el enzima glucosa – 6 fosfatasa, la obtención de
glucosa libre se puede hacer a partir de la glucosa – 6P y también del
glucógeno. Este órgano utiliza la glucosa para enviarla a la sangre, no para su
propio abastecimiento.
En cualquier otro tipo de célula
no se puede enviar glucosa libre a la sangre.
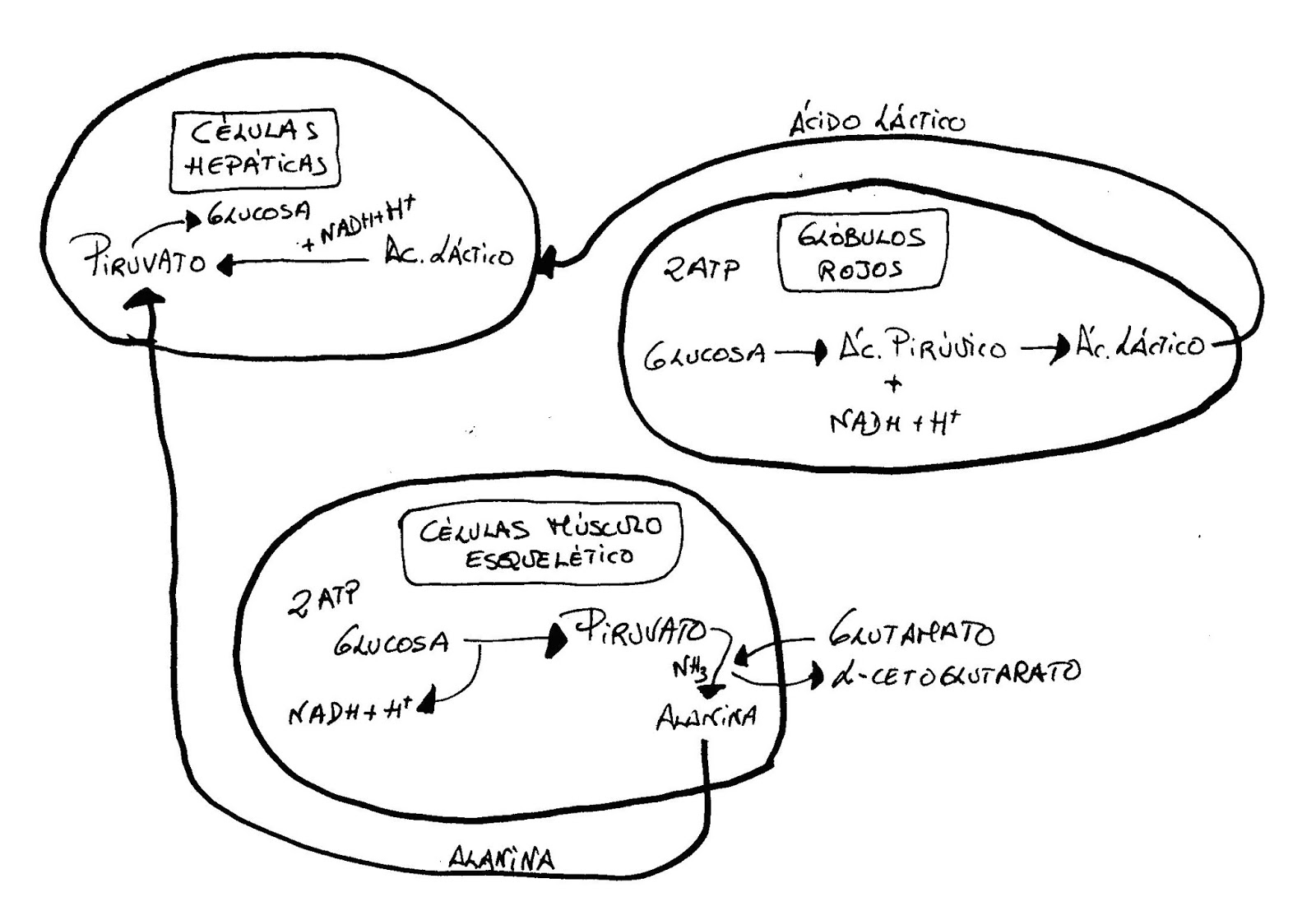
Las células hepáticas pueden
obtener Piruvato del metabolismo de otras células para producir glucosa.
Los glóbulos rojos;
mediante la glucolisis obtienen ácido Pirúvico y poder reductor, lo van a
transformar en ácido láctico que van a enviar al hígado. En el hígado se va a
transformar en piruvato (con liberación de NADH + H+) y el cual va a
ser transformado en glucosa.
En el músculo esquelético;
puede ocurrir lo mismo pero rentabilizando todavía más el transporte. Al
trabajar en anaerobiosis glucosa Þ piruvato y al mismo tiempo se pueden estar degradando
proteínas normalmente, experimentando procesos de desaminación obteniéndose
glutamato que reacciona con el piruvato que transforma en alanina por
transferencia de un grupo amino del glutamato Þ ά-cetoglutarato.
La alanina puede ir ahora a la
sangre sin transportador y luego al hígado. El poder reductor queda en la
célula del músculo esquelético siendo muy rentable para la célula este
transporte.
El proceso de gluconeogénesis se
activa siempre que hay sustratos disponibles. En ocasiones el músculo y tejido
periféricos producen estos sustratos y los envían al hígado para liberarse de
ellos y el hígado los utiliza para fabricar glucosa.
Si los depósitos de glucosa no
están cubiertos, el hígado puede usar estos sustratos para elaborar glucosa,
los va a almacenar en forma de glucógeno aunque no haya necesidad.
Con la cesión de ácido láctico al
hígado no sólo se obtiene piruvato sino que también se obtiene poder reductor.
En las células musculares hay una
gran masa proteica que sufre procesos de recambio, hidrolizándose de
aminoácidos y que tienden a transaminarse en una sola clase de aminoácidos;
L-glutamato. Además en el músculo esquelético se están realizando procesos de
glucolisis por lo que se genera piruvato, pudiéndose reducir ahí mismo en
anaerobiosis, reduciéndose a Lactato Þ Hígado (por la sangre).
Pero en otras ocasiones no se
transforma en lactato y el poder reductor se utiliza en la cadena de transporte
electrónico y el piruvato reacciona con el L-glutmato.
El L-glutamato al desaminarse se
transforma en ά-cetoácido (el ά-cetoglutarato) y el grupo amino lo envía al
piruvato que al servir un ά-cetoácido se transforma en un ácido, la alanina
(reacción de transaminación).
El ά-cetoglutarato puede ir
directamente al Ciclo de Krebs y el poder reductor a la cadena de transporte
electrónico siendo nada costoso la transferencia de la alanina al hígado. En
las células hepáticas, a la alanina le ocurre lo mismo que en las células del
músculo pero al revés.
Los ácidos grasos por su
degradación (β-oxidación), nunca podrán ser usados para la gluconeogénesis,
pero aportan poder reductor, que sí va a influir.
Cuando se produce la
gluconeogénesis en hígado, hay una retirada masiva de Oxalacetato del Ciclo de
Krebs de forma que el Acetil-CoA que llega al hígado tendrá que ser llevado por
otra vía para la obtención de otras moléculas combustibles, los cuerpos
Cetónicos.
Si en general se produce en el
cuerpo falta de glucosa y energía se movilizan las grasas del tejido adiposo
para poder abastecer estas necesidades.
El alcohol inhibe la
gluconeogénesis por lo que cuando un cuerpo está muy frío no hay que darle
alcohol para que se caliente, debe dársele glucosa. Cuando el alcohol entra en
el cuerpo se deshidrogena el etanol y produce poder reductor, desplazándose la
reacción piruvato →→ lactato hacia la derecha, habiendo muy poco sustrato
(piruvato) para fabricar glucosa que suele estar en niveles muy bajos en
estados de congelación. La sangre no llegará con suficiente glucosa, afectando
al cerebro y a los demás órganos. Además con el alcohol, si la glucemia ya
estaba baja, se produciría vasodilatación con lo que se perdería todavía más
calor aunque hubiera en un principio una sensación de calor superficial.
La alanina reaccionará con el
ά-cetoglutarato proveniente del Ciclo de Krebs para generarse piruvato y
L-glutamato que están dentro de las mitocondrias hepáticas.
Para construir glucosa a partir
de este piruvato, el hígado debe aportar poder reductor para poder transformarla, diferenciándose así
la llegada de lactato del glóbulo rojo y alanina del músculo.
El L-glutamato es forma de
amoníaco se transforma en ά-cetoglutarato. Este amoníaco es muy tóxico por lo
que el hígado lo tiene que neutralizar fabricando una molécula de urea en el
Ciclo de la Urea, es un proceso muy costoso energéticamente.
El músculo mejora su situación
pero el hígado tiene trabajo extra. Cualquier vía que aporte los intermediarios
del Ciclo de Krebs está promoviendo la gluconeogénesis:
Hidrólisis Desamina
GLUTAMINA----------------GLUTAMATO--------------------- ά-CETOGLUTARATO
Por lo que, la glutamina también
es precursor de la gluconeogénesis.
El ASPARTATO se considera
glucogénico (también la glutamina) porque son sustratos para la
gluconeogénesis. Los cetogénicos no darán lugar a la gluconeogénesis. Otro
precursor es el GLICEROL, producto de la degradación de los TAG.
Mediante pocas transformaciones sencillas genera intermediarios de la
gluconeogénesis

Dependiendo del momento
metabólico el glicerol pude ser usado para la glucolisis (energía) o para la
gluconeogénesis.
No hay comentarios:
Publicar un comentario