
Ladies and Gentlemen enfermer@s !!! un nuevo tema abordamos sobre bioquímica. Atención, puede ser muy probable que caiga también en el examen debido a su gran importancia en el metabolismo humano.
CICLO DEL ÁCIDO CÍTRICO o CICLO DE KREBS
Es una ruta metabólica central
que ocurre en la matriz mitocondrial donde se encuentran los enzimas que lo
catalizan. Es una ruta crucial que funciona siempre. Puede ser una vía tanto
para la síntesis como para la degradación. Se transforman muchas moléculas de
forma simultánea.
El Acetil-CoA puede seguir esta
vía, que es una ruta anfibólica cíclica que se usa en el metabolismo en procesos
catabólicos y anabólicos. Por cada vuelta de ciclo podrá ingresar un grupo
Acetilo proveniente de la molécula de Acetil-CoA que es poseedor de 2 C que se oxidará a lo largo
del proceso (8 etapas). Por cada vuelta se pierden 2 C en forma de CO2,
que no tienen porqué ser los que han entrado en el acetilo, y se liberan 4
pares de H.
- 3 transportados por NAD →→ NADH + H+
- 1 transportado por FAD →→ FADH2

ETAPAS
1-
CONDENSACIÓN DEL OXALACETATO Y ACETIL
El
Oxalacetato es una estructura de 4
C que como resultado de su condensación con el Acetil da
lugar al CITRATO. Está regulado por el enzima CITRATO SINTETASA, enzima
controlado por la concentración del producto final (* punto de control).
En
este momento se libera la molécula de CoA para que pueda tener lugar la
condensación. El CoA puede ser usado en otra descarboxilación oxidativa del
Ácido Pirúvico.
2-
CONVERSIÓN DEL CITRATO EN ISOCITRATO
El
Citrato es un intermediario que puede abandonar el Ciclo y salir de la
mitocondria, pasar al citoplasma y ser utilizado para la síntesis de ácidos
grasos. Si permanece en el ciclo tiene que transformarse en una estructura que
pueda oxidarse y no se rompa tras varias etapas se transforma en ISOCITRATO
(enzima, ACONITASA).
Si
se usa para la síntesis de ácidos grasos, como se necesita Acetil-CoA y este no
puede abandonar la mitocondria en esta forma, lo que va a hacer es desdoblarse
en: Oxalacetato + grupo Acetil-CoA.
3- ISOCITRATO a ά-CETOGLUTARATO
Reacción
catalizada por la isocitrato deshidrogenasa.
El
Isocitrato posee 6 C
y puede deshidrogenarse sin romperse mediante el enxima Isocitrato
deshidrogenasa, el isocitrato se transforma en ά-CETOGLUTARATO (5 C ).
Se
genera una molécula de poder reductor. Es una reacción muy importantes. Se
pierde un ion hidruro que es aceptado por un coenzima y se libera un carbono en
forma de CO2 que podrá abandonar el citoplasma y ser utilizado como
un deshecho o ser usado para futuras descarboxilaciones.
El
coenzima que suele actuar es el NAD pero también puede ser el NADP.
El NADH + H+
puede regenerarse en la cadena de transporte electrónico donde se libera
energía para la síntesis de otras moléculas. Ocurre en la mitocondria. El NADPH2
no puede regenerarse en la cadena de transporte electrónico pero sí puede
liberar energía en un proceso de síntesis.
Este proceso es de control. El
enzima que loruela está controlado por la concentración de NADH + H+ ya que elevadas
concentraciones provocan la reducción de la actividad enzimática. Al
ά-cetoglutarato se le puede añadir un grupo amino dando lugar a un aminoácido
L-Glutamato. Pero por desaminación de glutamatos también se obtiene ά-cetoglutarato.
4. ά-CETOGLUTARATO CoA a SUCCINIL-CoA
Funcionamiento regulado por
aumento de la [ATP], reacción catalizada por la
alfa cetoglutarato deshidrogenasa.
El Succinil-CoA tiene 4 C . esta reacción está
catalizada por un complejo enzimático (3 enzimas y 5 coenzimas), llamado
ά-cetoglutarato deshidrogenasa y su funcionamiento es regulado por elevadas
concentraciones de ATP.
Una deficiencia en las vitaminas
provocaría la alteración del complejo. Se produce la pérdida de un átomo de C
en forma de CO2 y la deshidogenación
del sustrato que son aceptados por NAD. Se obtiene la 2ª molécula de poder
reductor. También hay el aporte de un CoA que queda integrado en el Pf.
5. SUCCINIL-CoA a SUCCINATO
Reacción catalizada por la
succinil CoA sintetasa, aquí es el único lugar donde se libera energía en forma
de GDP. (GDP + P = GTP, transfiere sus energía al ADP quedando ATP y de nuevo
ADP)
El Succinil-CoA es una molécula
muy inestable con exceso de energía que tiende a liberar para formar una
molécula energética.
El succinil libera parte de su
energía para la síntesis de una molécula de GTP que va a ceder su grupo P al
ADP dando lugar al ATP y el se transforma en GDP.
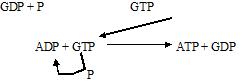
Esta es la primera y única
molécula que se produce en el ciclo. Durante este proceso también se
libera la molécula de CoA que puede ser
utilizada de nuevo.
6. SUCCINATO a FUMARATO
Reacción catalizada por el
succinato deshidrogenasa.
El succinato es oxidado a
Fumarato. El coenzima que actúa es el FAD aceptando los H2 del
succinato. El FAD permite una conexión directa con la cadena de transporte
electrónico.
1 NAD + H+ ------------------------------- 3 ATP
1 FADH2 -----------------------------------2 ATP
7. FUMARATO a MALATO
El Fumarato a través del enzima FUMARASA da lugar a
Malato
8. MALATO a OXALACETATO
Reacción catalizada por la malato
deshidrogenasa. (se cierra el ciclo)
El Malato experimenta la última
deshidrogenación del ciclo y se convierte en Oxalacetato (reacción reversible).
El Ciclo de Krebs o del Ácido
Cítrico tiene lugar en el sentido de las agujas del reloj. Es un proceso
aerobio.
BALANCE TOTAL:
2
moléculas de CO2 1 FADH2
3 moléculas de NADH+H+ 1 ATP

- La enzima que cataliza el succinil-CoA es la succinil CoA sintetasa
- La que cataliza el succinato es la sucinato deshidrogenasa
- La que cataliza el malato es la malato deshidrogenasa
Los intermediarios de este ciclo
pueden ser administrados a través de otras rutas metabólicas. Una de ellas es el aumento de la concentración de
Oxalacetato que se necesita para aceptar grandes cantidades de Acetil-CoA por
una necesidad de energía inmediata. El Acetil-CoA puede proceder de la glucosa
(piruvato) o de los ácidos grasos.
El aumento de la concentración de
oxalacetato se consigue mediante dos rutas anapleróticas, partiendo de
piruvato.
1- El obtenido en la célula
procedente de la glucolisis.
El piruvato es transformado en oxalacetato y es
catalizado por el enzima piruvato-carboxilasa. El enzima
piruvato-deshidrogenasa actúa como modulador del piruvato-carboxilasa.
El piruvato posee 3 C y el Oxalacetato posee 4 C . El C que le falta proviene
del CO2 liberado en el ciclo del Ácido Cítrico, que reacciona con el
piruvato y a través de distintas etapas y por acción del enzima da lugar al
Oxalacetato.
2- Es menos utilizada.
El
piruvato se transforma en Malato (4C )
que después pasa a oxalacetato.

Para ello es necesario una
carboxilación (CO2 del Ciclo de Krebs) y un aporte de poder
reductor (NAD/NADP) que también se
obtiene del Ciclo de Krebs y que no se podía usar en la cadena de transporte
electrónico. El NADP se regenera. El Malato una vez obtenido experimenta
deshidrogenación y libera poder reductor.
Cuando una célula hepática quiere sintetizar glucosa es necesario la
presencia de una concentración elevada de oxalacetato. Este oxalacetato puede
provenir del Ciclo de Krebs, pero no tiene sentido porque no sería rentable,
pero puede obtener piruvato de otras células como la célula del músculo
esquelético. El piruvato de esta célula se transforma en lactato que, por la
sangre, llega a la célula hepática donde se transforma en piruvato que da lugar
al oxalacetato necesario para la gluconeogénesis.

No hay comentarios:
Publicar un comentario